
The origin of mankind
THE ORIGIN OF MANKIND
Francesco Callea
Catholic University of Mwanza, Tanzania
Creation and Evolution theories share two major features: uniqueness and variability of human kind. This approach is new in that bypasses the age-old problem about their conflicting relationship.
In the Bible (Genesis 1 and2) there are at least 4 examples showing how Creation and Evolution theories concerning the origin of mankind share the above two major properties: Babel’s Tower, the creation of man, the Great Flood and the creation of Eva.
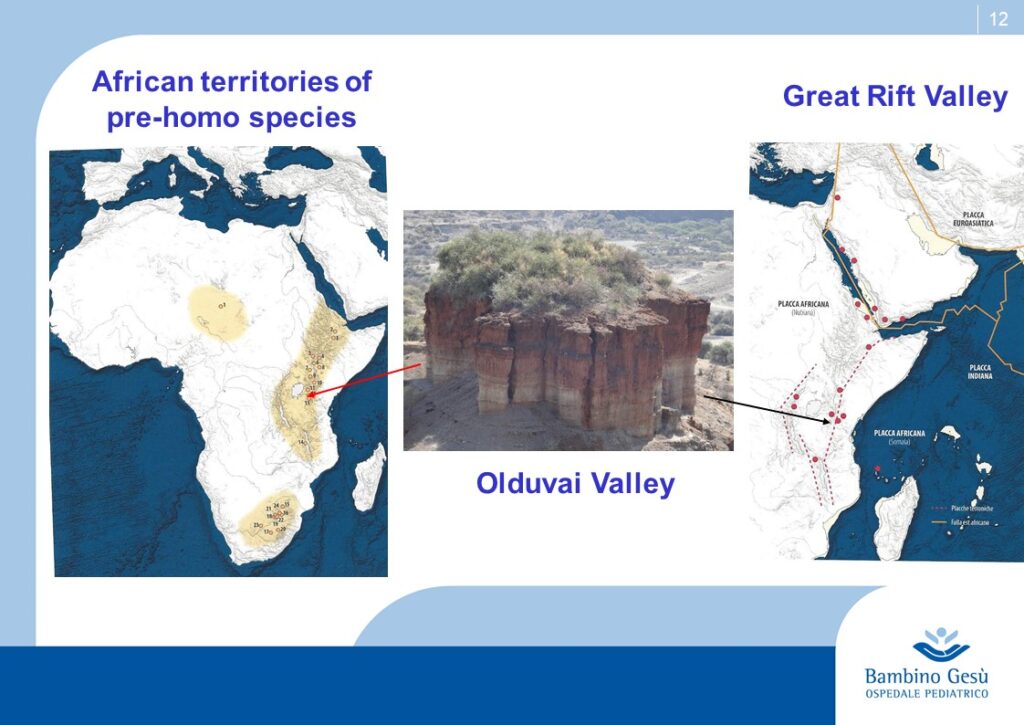
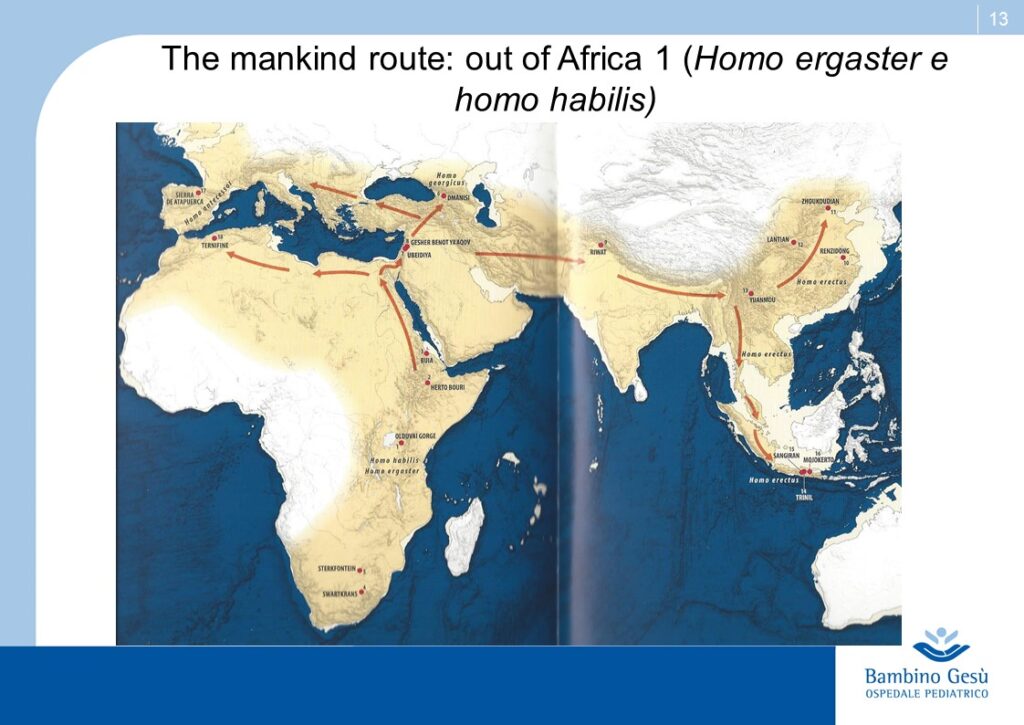
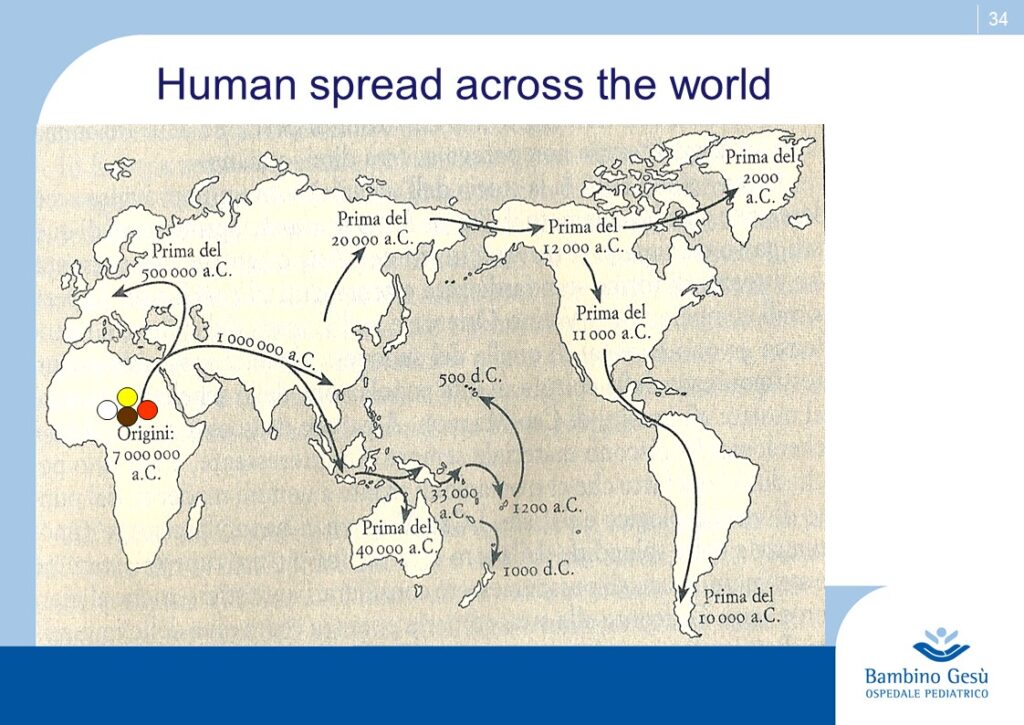
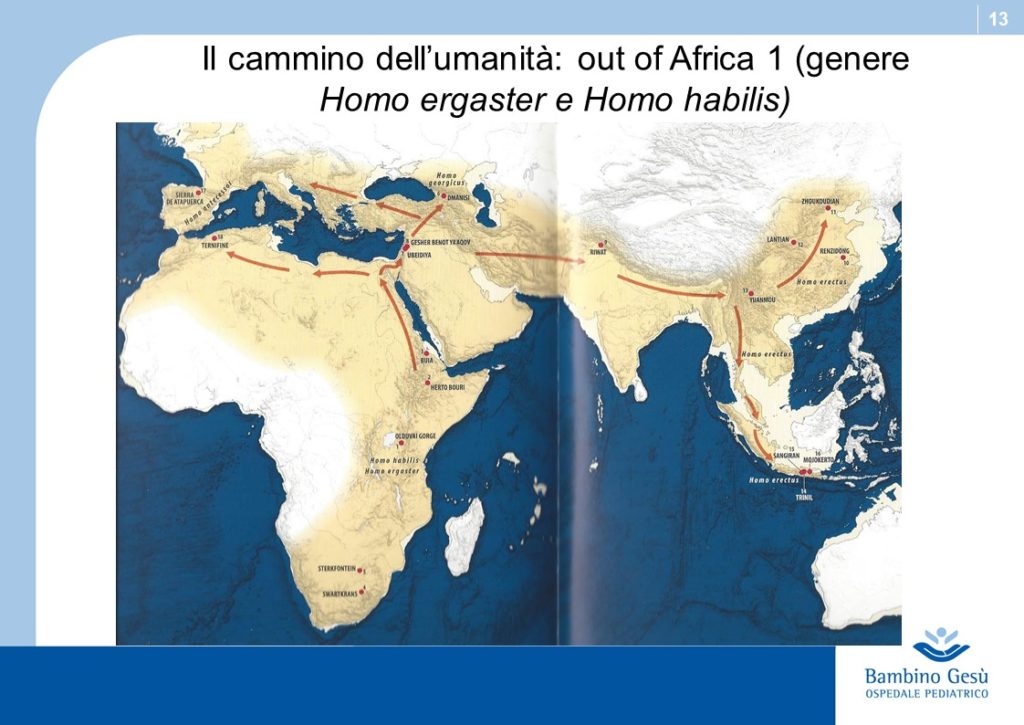
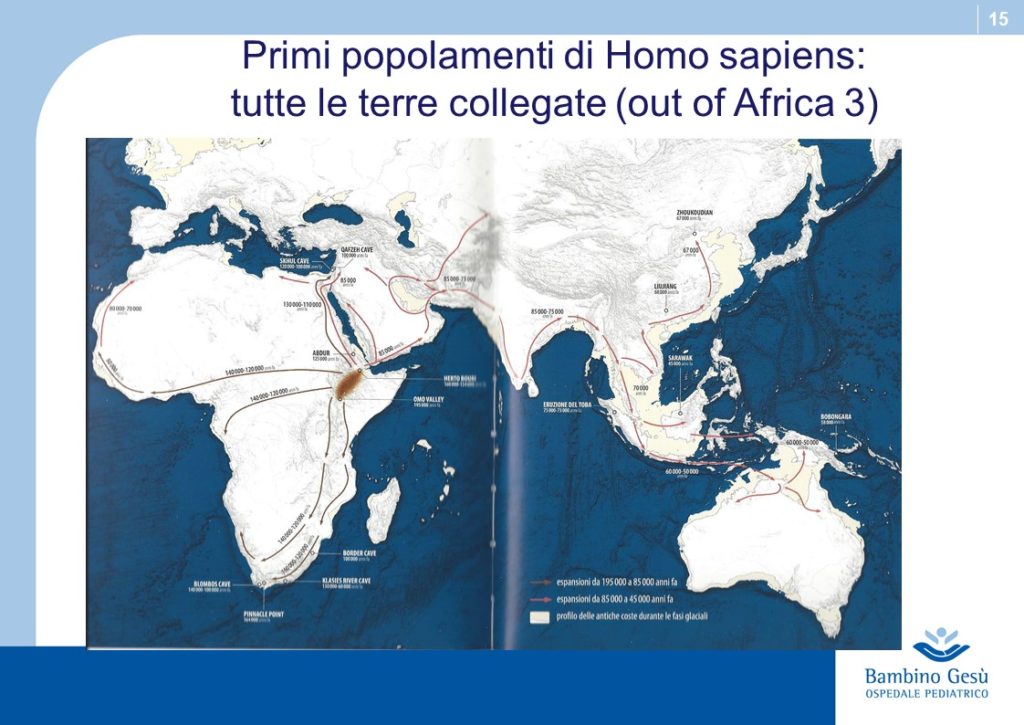
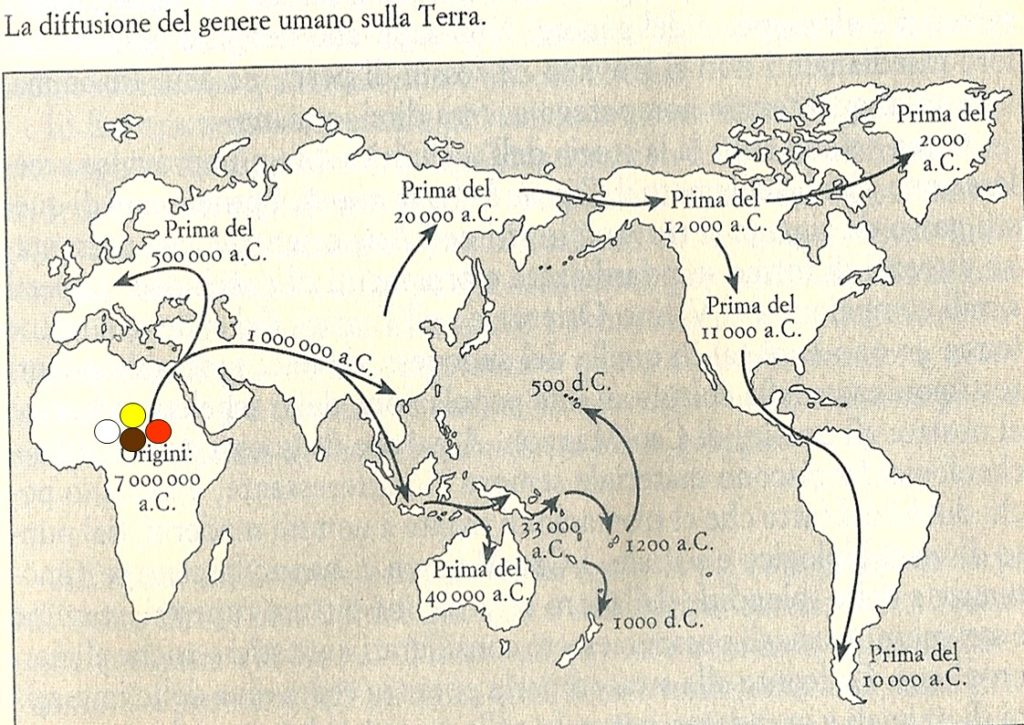
Evolution theories are based upon the results of paleontologic, anthropologic, ethnic, linguistic and genetic studies. Along Adam and Eva’s traces, the molecular clock and genoma studies, the genealogic tree of human kindship has been reconstructed (1) . The comparative studies have confirmed that the first traces of humans have appeared in the Northern Tanzania and that the migration process known as ”out of Africa” has started from that area of the world for further colonization of all lands of the planet.
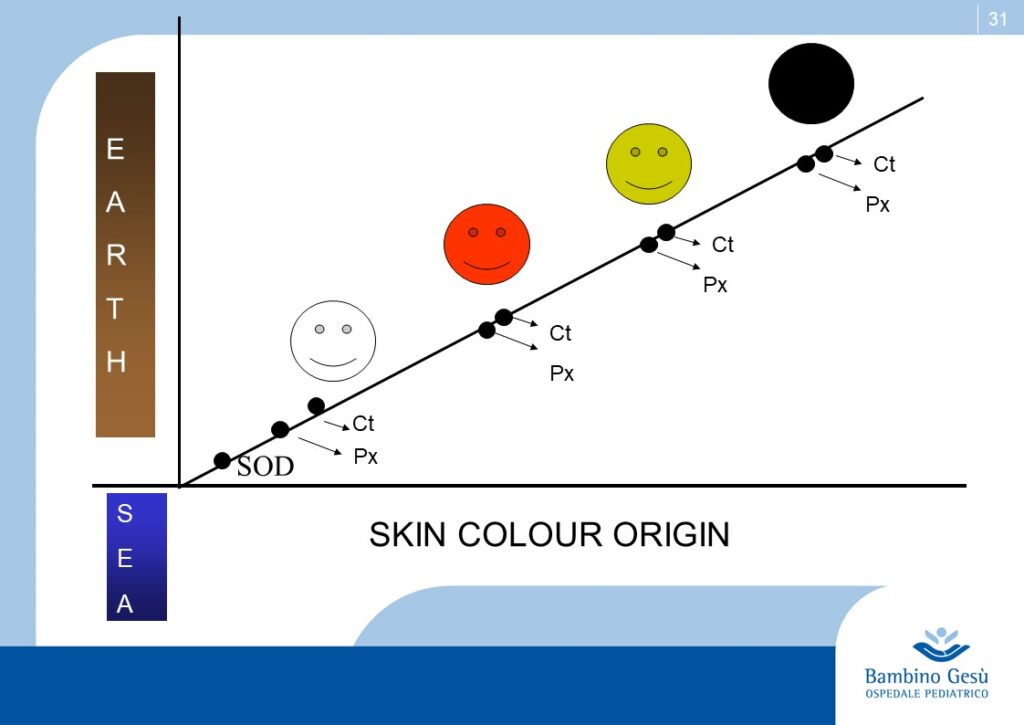
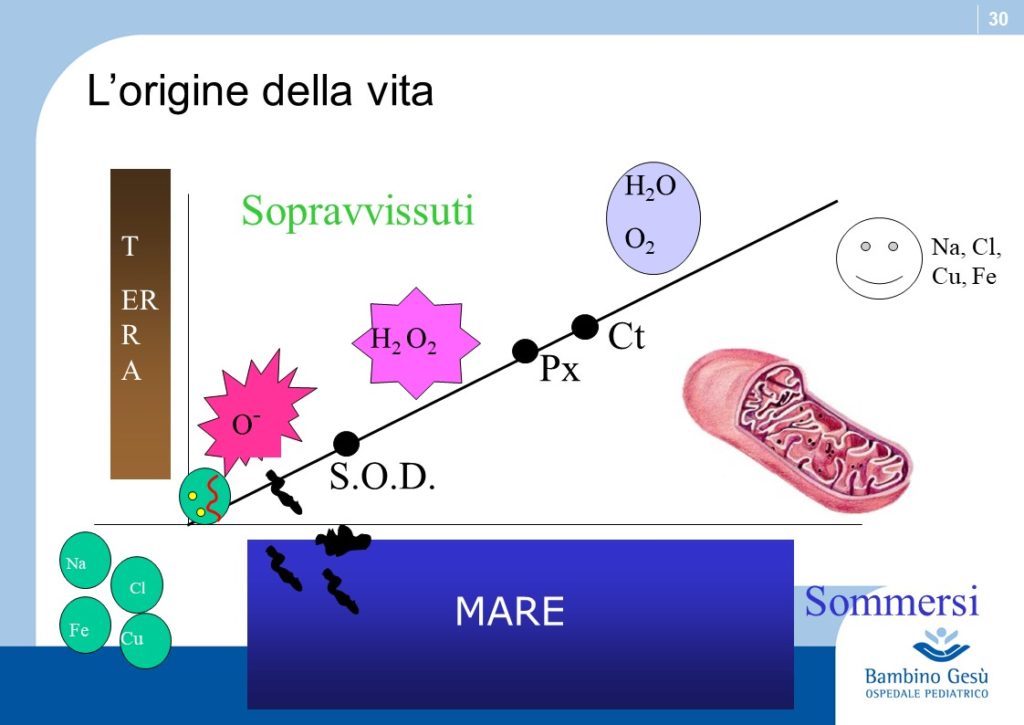
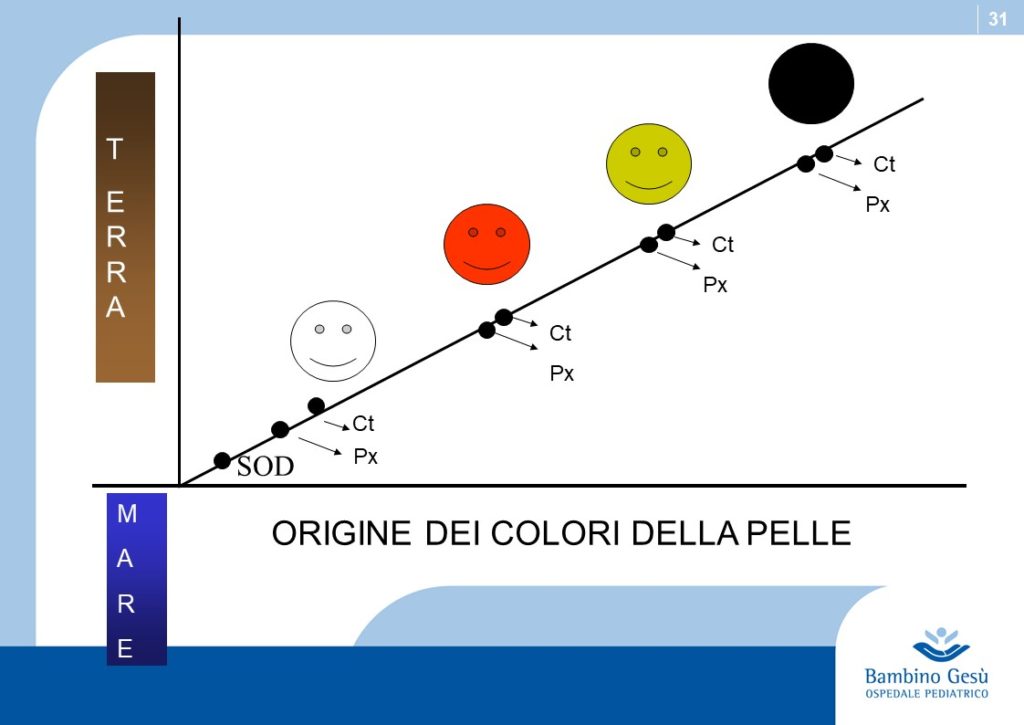
Skin color is one of the most conspicuous ways in which humans vary and has been widely used to define human races. Recent studies have definitely proven that the variations in skin color are adaptive and related to the regulation of ultraviolet (UV) radiation penetration in the integument and its direct and indirect effect on good health (2)
The earliest members of the mankind lineage probably had a mostly unpigmented or lightly pigmented integument covered by dark black hairs, similar to that of the modern chimpanzee (3). The evolution of a naked, darkly pigmented skin occurred early in the evolution of the genus Homo. A dark epidermis protected sweat glands from UV-induced injury, thus insuring the integrity of thermoregulation.
Of greater significance in individual reproductive success was that highly melanized skin protected against UV-induced proteolysis of folate, a metabolite that is essential for normal development of the embryonic neural tube and spermatogenesis.
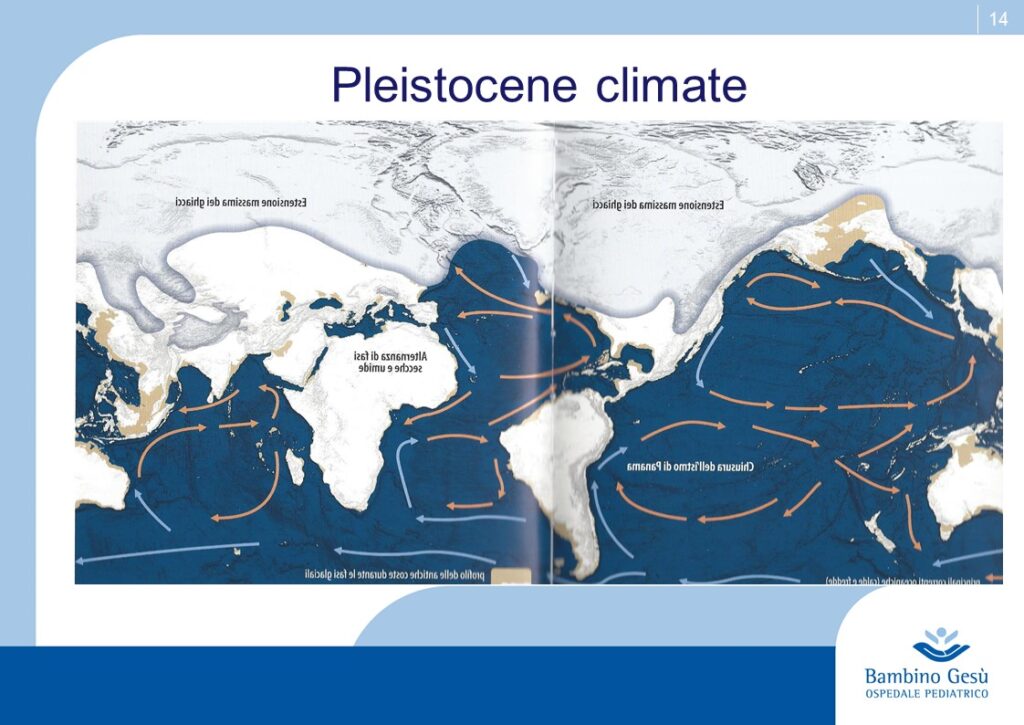
As hominides migrated out of Africa, varying degrees of depigmentation evolved in order to permit UV-induced synthesis of previtamin D3. Generally speaking the impact of UV-eradiation on skin increases by latitude and decreases by altitude. Recent observations explain the apparent exceptions to the general rule, like the Ituin group in Eskimo population. In general, females require a lighter color of the skin to synthesize more amounts of Vitamin 3, necessary during pregnancy and location (3)
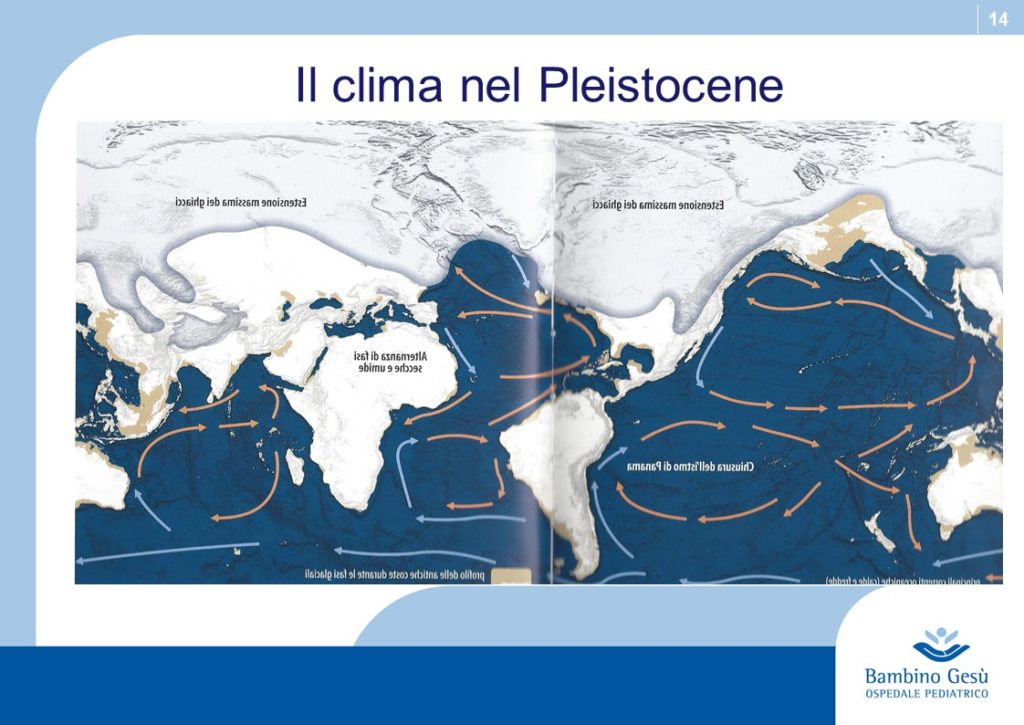
Thus skin coloration in humans is adaptive and labile. Skin pigmentation levels have changed more than once in human evolution (similar to the alternation of glaciation and desertification). Because of that, the skin coloration should not be considered of value in determining phylogenetic relationship among modern human groups.
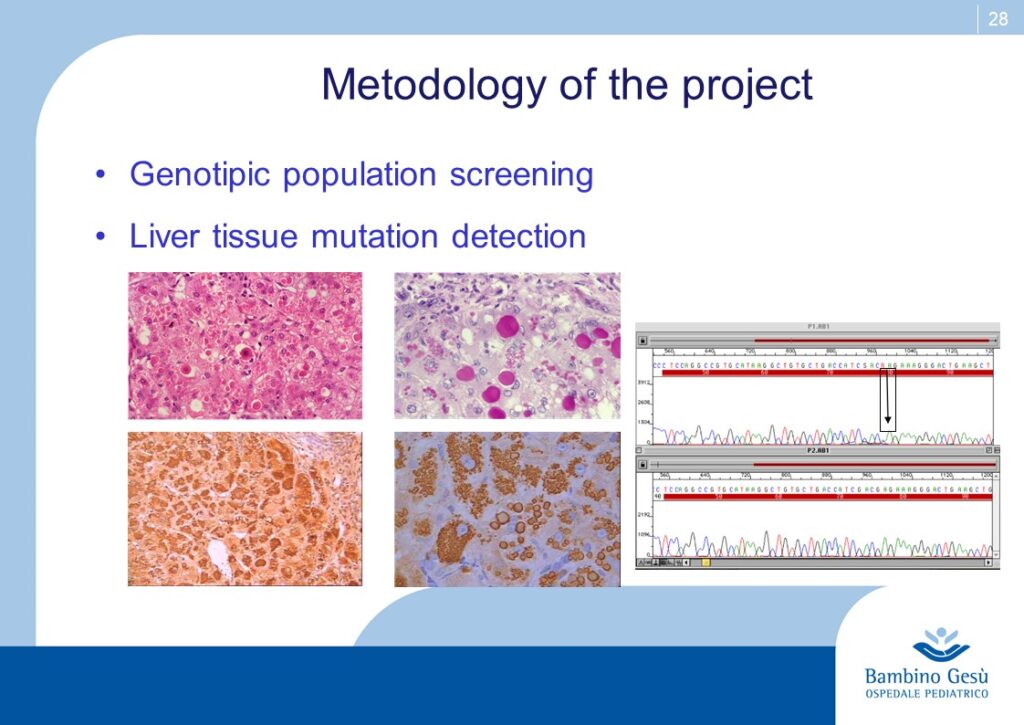
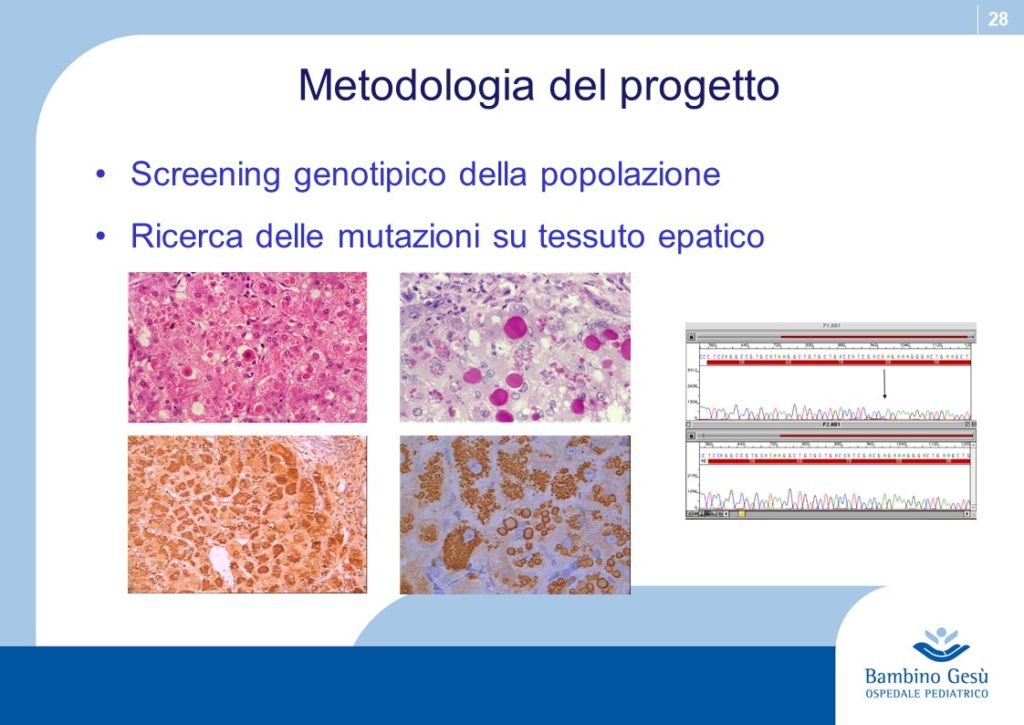
An ongoing research project supported by two Italian ONLUS Associations (“Friends Raising” and “Alpha-1-antiyrypsin”) is aimed to search in the population of Northern Tanzania the pathogenetic mutations that have been detected in Europe, Asia, America, Oceania and to verify their role ibn the development of cryptogenic cirrhosis and HCC (4) that carry a high incidence in that area of the world. The background of the project is based upon two robust points: the “out of Africa” migration process and upon the finding of pathogenetic mutations in black Americans (Afro-Americans) (5) who have moved from the black Africa to America in more recent years for reason other than the original “out of Africa”. The project sounds as Alpha-1-antitrypsin would return back home.
References
- Cann GR et al. Genealogic tree of human population, Nature 1987 in Homo Sapiens. Cavalli Sforza LL et Al “The story of Human Diversitry” 2013
- Diamond J.. “Geography and skin colour”. Nature 2005;435:383-384
- Jablonski NG et al. “ The evolution of human skin coloration”. J Hum Evol 2000,39:57-106
- Callea F. et al. “Mineralization of alpha-1-antitrypsin inclusion bodies in Mmalton alpha-1-antitrypsin deficiency”. Orphanet Journal Rare Disease 2018;13: 79
- Pierce JA et al. “Antitrypsin phenotypes in St. Louis”. JAMA 1975;2238:609-617
Congresso Italiano di Pediatria

Domani – n° 49, Marzo 2018


Ethnic groups Origin. Wolrdwide geographic distribution of AATD.










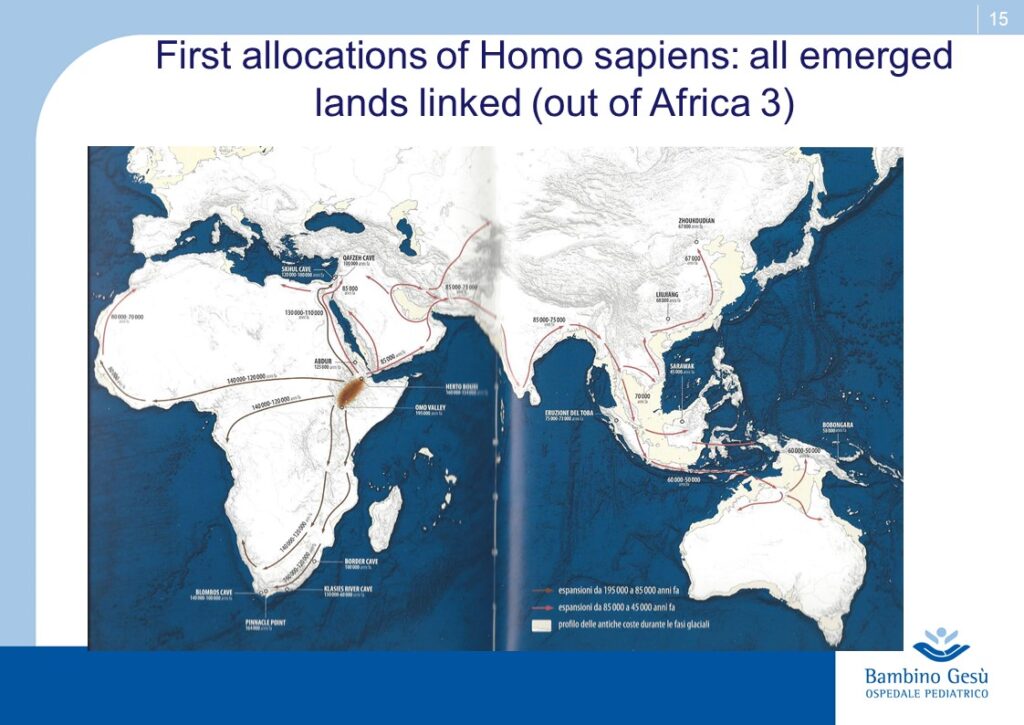
















L’origine delle razze. Distribuzione geografica dell’α1-anti-tripsina nel mondo
























La Provincia Cosentina

Friends Raising e Alfa1AT



